
Pea Peptide Study, Leaky Gut and Lipedema
As a former lab rat, I have so much interesting stuff to share with you.

Flashback to this day six years ago. I was a lab rat, and I loved it. It was everything I had ever dreamed of.

In this experiment, I worked on pea peptides as part of a research program called FoodProFuture, which explored plant proteins. There were so many fascinating studies I could have done within the program. I was initially drawn to investigating antinutrients in food, but I ended up choosing a cell-based study to look at inflammatory responses and to see how much of the material was taken up through the intestine. As you can see in the photo, I’m testing whether the model is working as intended.
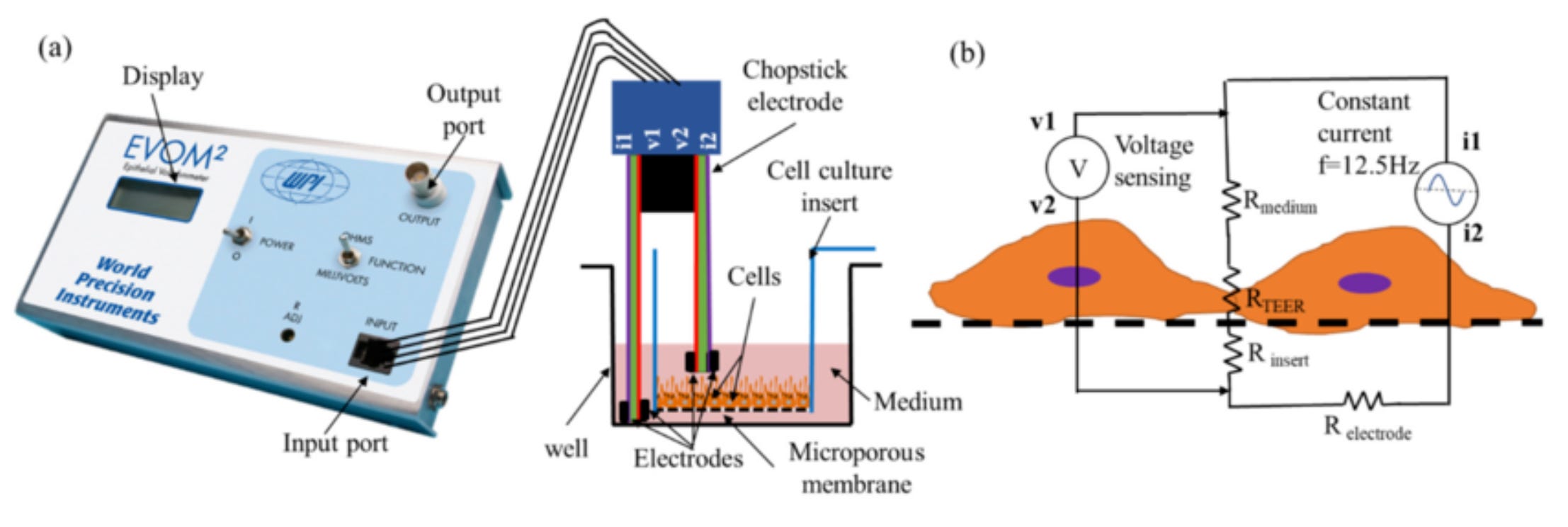
I grew Caco-2 cells (in vitro) to form an intestinal-like barrier, then added the pea peptides, applied an electrical pulse to measure transepithelial electrical resistance, and analyzed the material on the other side to see whether I could detect the pea protein.
Caco-2 cells are a human colorectal adenocarcinoma cell line that spontaneously differentiates in culture to mimic the small intestinal epithelium. They are widely used as an in vitro model for studying drug absorption, intestinal permeability, and nutrient transport due to their ability to form polarized, tight-junction-containing monolayers. They are commonly used in pharmaceutical research.

It was genuinely fascinating. I learned so much about the components between the cells that regulate what gets through and what does not.
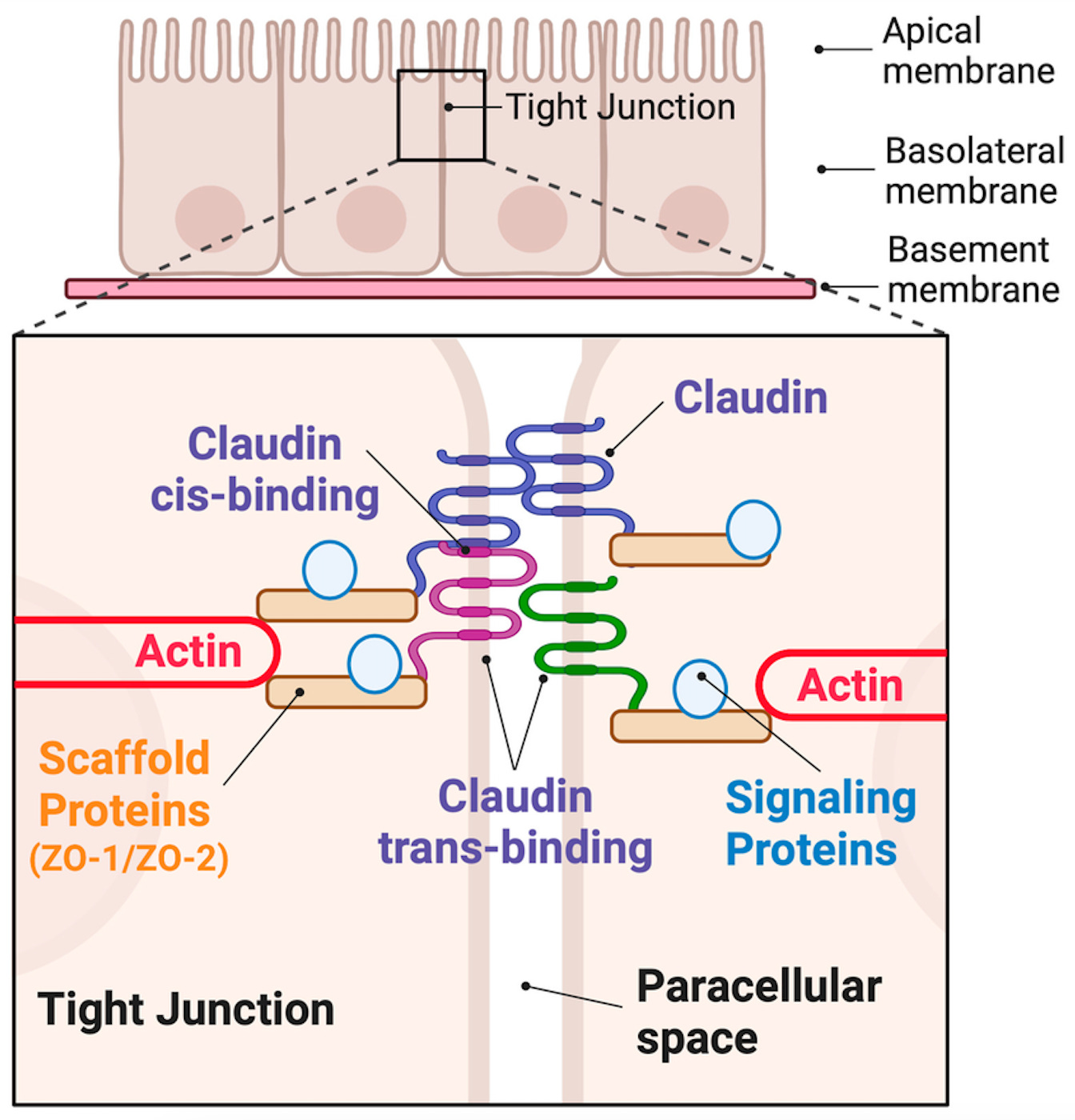
What made these in vitro studies on Caco-2 cells so incredibly exciting was learning about the complex dance between the cells in the intestinal barrier, especially the tight junctions that regulate what is allowed to pass through the paracellular space.
Tight junction proteins such as claudins and ZO-1 form a highly regulated seal between epithelial cells and prevent uncontrolled leakage of molecules. In the Caco-2 model, one could directly observe how transepithelial electrical resistance (TEER) droppes when the junctions were disrupted by inflammatory stimuli or bioactive compounds, effectively opening the paracellular pathway. Claudins interact both within the same cell membrane and across adjacent cells, supported by the actin cytoskeleton and scaffold proteins such as ZO-2. Together, they regulate selective permeability in a highly dynamic manner.

One of the most fascinating aspects was how interconnected the cells are. When one cell weakens, it affects its neighbors through altered claudin expression and reduced localization of ZO-1 at the membrane. In our experiments, lipopolysaccharide-induced inflammation reduced TEER and increased permeability to markers such as FITC-dextran. At the same time, certain compounds, including probiotic-derived metabolites or plant bioactives, were able to restore barrier integrity by upregulating tight junction proteins. The paracellular transport increased under stress, but signaling pathways such as AMPK and NF-κB were also activated in attempts to repair the barrier.
Seeing ZO-1 and occludin disappear from the cell borders under inflammatory stress, and then reappear after treatment, made it clear how dynamic and responsive the intestinal epithelium truly is. It changed my understanding of how nutrients, inflammatory signals, and dietary components can modulate the entire barrier system. This is deeply relevant to what is often described as increased intestinal permeability, or “leaky gut.”
This connects directly to lipedema. Just as we measured individual variations in TEER responses in the Caco-2 model, intestinal barrier integrity varies from person to person. Differences in claudin expression, ZO-1 levels, cytoskeletal organization, and microbiome composition influence how nutrients and endotoxins are absorbed. This may help explain why there is no universal diet that works for everyone living with lipedema.
If you value what you find here and appreciate the work CarinaW is doing for the lipedema community, consider becoming a subscriber. Your support is what makes this platform sustainable. It allows independent, science-based content to continue growing, reaching more women, and deepening the collective understanding of lipedema.
In individuals with lipedema, tight junction function may be compromised in some cases, increasing paracellular permeability and allowing endotoxins such as lipopolysaccharide to translocate into circulation before nutrients are fully absorbed via transcellular pathways. This can contribute to systemic inflammation, which may exacerbate inflammatory processes in subcutaneous adipose tissue. Because claudin expression patterns, microbiome diversity, and inflammatory signaling differ between individuals, responses to carbohydrates, fats, or fiber can vary significantly.
Research in metabolic disease shows that dysbiosis can weaken tight junction integrity, reduce production of short-chain fatty acids such as butyrate, and increase circulating endotoxin levels. Butyrate normally strengthens ZO-1 expression and supports barrier integrity. Reduced microbial diversity and increased pro-inflammatory bacterial species have been observed in obesity and metabolic syndrome, conditions that share some inflammatory characteristics with lipedema. Hormonal and biochemical alterations in lipedema may further amplify this vulnerability.

Increased intestinal permeability occurs when tight junction proteins such as claudins are disrupted, leading to elevated paracellular flux. In Caco-2 cells, TEER drops after LPS exposure, mirroring what is seen in dysbiosis-related barrier dysfunction. Endotoxins can then translocate into the bloodstream, activating systemic inflammatory pathways that may worsen the tender, fluid-rich adipose tissue characteristic of lipedema.

Microbiome alterations in lipedema are still under investigation, but patterns seen in obesity-related inflammation provide important clues. Dysbiosis reduces mucin production, increases LPS-producing bacteria, and impairs epithelial repair mechanisms. Human studies link microbiome shifts to metabolic endotoxemia, while in vitro studies show that probiotics and bioactive compounds can restore tight junction integrity in epithelial models. For lipedema, this suggests that personalized strategies targeting gut barrier health and microbiome balance may be essential before expecting dietary interventions alone to improve symptoms.
What the lab taught me was this: the intestinal barrier is not static. It is alive, adaptive, and profoundly individual. And that biological individuality may be one of the missing pieces in understanding why lipedema responds so differently from one woman to another.